1.
肉牛生產背景
1.1 影響牛隻生產的歷史特性
日本除了家豬與家鴨有野豬種 (Sus
suofa) 與野鴨種 (Anas sp.) 一併存在之外,就沒有本土自然生存的一般家畜祖先種。家畜如豬、牛和雞曾在繩文年代晚期 (-BC 500) 至彌生年代 (BC500-AD300) 引進日本。中國歷史書籍 (-AD250) 也記載,日本沒有牛馬及羊。有關豬和雞,由於該書籍中並沒有發現任何的記載,牠們當時可能存在的事實是不能給予否定的。
在第七世紀有數種書中曾提到牛乳。(英吉西克,927)在平安年代的著作中,描述過乳製品: 酥,「蘇,“so”」。據推測係一種煉乳,是為醫療目的使用。書中還提到貢獻“蘇”給政府,可是這樣的慣例在 12 世紀初已經廢除。到西元 675 年政府禁止屠宰動物包括牛、馬、狗、猴和雞等,而且在西元 742 年又重新禁止屠宰牛馬。有這種殺生禁令暗示人們在這段時期有過食肉的事。
從禁令以後,肉類和乳類的利用就變成愈來愈不普遍。雖然在西元 1700 年左右寫下的圖畫書上有乳製品的介紹,然而家畜主要用途是貨物運輸、農耕或軍用勞力如馬的役用,同時也有一些農業書籍介紹為肥料生產的飼養方式。
日本與其他大部分的國家相比,為實際食物的生產而飼養家畜的歷史,是極短的。這事實看來是由於下述兩種原因:其一,日本的氣候條件適合榖類耕作,而養牛唯一用途是幫助稻米耕種。其次,日本佛教長久以來佔有優勢,並且禁止食肉,特別是有四隻腳的動物。因此造成畜產品的利用並不普遍,一直到明治時代以前,特別在日本中部地區是如此。其實日本有肉類的消費,差不多只有 130 年而已,始於明治時代初期,可是食肉大眾化還是最近 30 年的事。所以,可以說在 1950 年中期以前,日本的牛隻未曾接受為產乳與產肉而進行的技術上改良。後來在 1950 年日本制定了家畜改良及增殖法。該項法律規定政府必須確定改良及增殖家畜的目標,其詳細說明為〝農林水產大臣應依家畜種類訂定具體的目標,該家畜種類包括牛、馬、綿羊、山羊、豬及其他根據法令所規定的家畜在內並且應公告改良繁殖的每一項目標〞。
在明治時代,已有很多的外國牛被引進日本,並在政府領導下,起初的時候是廣泛與本地的牛隻雜交。經過該階段,日本牛隻的基因庫得以稀釋且大量擴充。在過了開始時一陣狂亂的雜交利用後,各牛種的育種家就著手自己牛隻品種改良,不再利用雜交。此舉又得到各縣的推動,因此各縣得以育成如今所見的日本牛種,具有獨一無二的特徵。由於各地區從事引進與繁殖改良結果,造成日本的本地牛純種大量消失,僅剩下兩個島保存:山口縣的見島和鹿兒島縣的口之島(如圖一)即是。
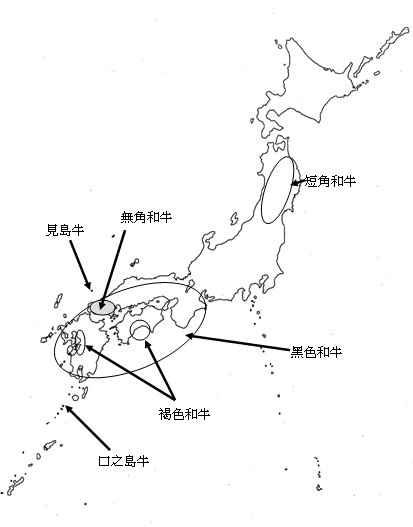
圖 1. 日本的本地牛品種和族群的原產地。
1.2 家畜在日本日常生活中的地位
有很多的傳統活動與各種家畜有關係,而且現在依然持續著,特別是與牛和馬有關的。這些包括:和霊神社的“牛鬼 (Usioni)”,太秦 (Uzumasa)的牛節及日本農村田間工作有關的宗教儀式 (津田, 2001)。在這些活動中,活著的家畜扮演帶領角色,但卻很少出現未改良的本地種家畜。現今,改良過的品種的確在這些傳統的活動和節日中被使用(照片 1)。

照片 1. 日本古代流行節日所用的牛。
從畜禽生產說起,如今所從事的是大規模飼養;平常生活中對家畜的存在與認識已逐漸消失。畜禽中只有少數的種被育成具有特殊的用途,例如:鬥牛用的牛,及鬥雞(軍雞,Shamo)、尾長雞 (Onagadori)和長鳴雞(Naganakidori)等作為鬥雞和玩賞雞。此外,在沖繩縣以山羊肉與豬肉作成的菜餚,和在秋田縣使用比內土雞做出的切棉團子―煮鍋 (Kiritanpo-nabe) 。可以說,傳統的佳餚類型利用日本傳統的品種做出。今日的日本平常生活中,已經熟見的是非本地的品種。除了雞之外,雖然佳餚和品種由前代傳給後代,卻看不出導致本地家畜的保存。
2. 日本的本地牛品種概述
日本
2000 年肉牛總頭數為二百八十二萬四仟頭以及乳牛總頭數為一百七十六萬四仟頭。肉牛可以分為兩種基本的類型:一種是日本的本地肉牛,有一百七十萬頭;另一種是非本地肥育用乳牛,有四十六萬一千頭,以及雜種牛,有六十六萬三千頭。日本的本地肉牛品種即稱為和牛(Wagyu),共有四個品種:黑色和牛
(Japanese Black,
93.9%)、棕色和牛 (Japanese Brown, 4.2%)、無角和牛 (Japanese Poll,
少數)和短角和牛 (Japanese Shorthorn Cattle, 1.0 %)。各個品種均具有其歷史及明顯的特徵,擬在下段詳細加以說明。所有產乳的品種都是外來的,況且超過百分之九十九的乳牛是荷蘭牛,由兩種純本地牛種所生產的乳品,所佔的比例確是微乎其微(資料來源:
2000 年家畜改良有關統計資料)。
2.1 見島牛 (Mishima cattle),
(如照片 2)
![]()
照片 2. 見島牛(公)。
見島 (Mishima) 位於北緯 34° 46', 東經 131° 8' 的日本海,其面積為 7.8 平方公里。見島是一個面積小且為平地有限的島嶼,因此其稻田面積小且為梯田。見島牛身體大小為小型及性情溫順而適合小農。最早的官方紀錄指出, 1672 年有 350 頭牛因牛瘟而全部消滅,之後又有新的牛群被建立起來。在 1739 年有 433 頭牛的紀錄且明治維新(1869)前大約有 400 頭牛被飼養。在 1928 年時見島牛被認為黑色和牛的原型,而被指定為日本天然紀念物。在指定之後,見島牛一直被當作家畜飼養並且仍在原地保種。到 1961 年見島牛母牛群飼養頭數已超過 300 頭。此後飼養頭數急劇減少,到 1970 年代中期只剩下 33 頭母牛 (古川等, 1997),而後母牛頭數又逐漸增加,到 2002 年就接近於 100 頭。
見島牛是熟晚的品種,與現代的日本黑牛比較起來體軀狹窄。皮毛呈暗褐色而頭角比較小。成年母牛(
60個月)的平均身高、胸圍和體重,分別為 112.8 公分、152.1 公分和
261.1 公斤(原田,
1996)。
2.2 口之島 (再野生化)
牛 (Kuchinoshima feral
cattle), (如照片 3)
![]()
照片 3. 口之島牛。
口之島
(Kuchinoshima) 是位於北緯
29°58', 東經 129°55', 其面積為 13.3 平方公里,離九州南方
200 公里處,即在吐卡拉群島北端的位置上。 1727 年的紀錄指出口之島有馴化牛的存在
(富田,1996),然而林田和野澤 (1964) 提出該野生的牛該是
1918 到 1919 年間由鹿兒島引進牛隻的後代從放牧場地逃出來的。在
1999 年該島的口之島牛現有頭數是 44 到 66 頭。另外有一部分的口之島牛保存在鹿兒島大學和名古屋大學,在
2001 年其頭數分別為 20 頭和 24 頭。口之島牛的身體大小是比見島牛來的小。成年母牛的平均身高及胸圍各為
110 公分及 120 公分。毛色主要是黑色帶有白色斑點在腹部和/或在四肢,偶爾也會出現褐色毛色。
2.3 黑色和牛
(Japanese Black) (如照片 4)

照片 4. 黑色和牛(公)。
大部分的黑色和牛是經過雜交後育成並使該品種擁有現代體型。在江戶年代 (1600 - 1876) 中國地區已經選育數種雜交前的品系 (Tsuru),其用途主要為搬運木材以供熔化鐵沙製鐵,當做役用家畜利用。在 1867 年明治維新後,為了改善牛隻身體大小及產乳量,政府鼓勵人們引進外國品種與本地牛雜交。如表 1. 所示
表 1. 日本各縣進行的外國品種與本地牛雜交
|
現代的品種名稱 |
縣別 |
外國品種 |
|
黑色和牛 |
京都 |
褐色瑞斯牛 |
|
|
兵庫 |
短角牛,德盆牛,褐色牛 |
|
|
|
瑞士牛 |
|
|
岡山 |
短角牛,德盆牛 |
|
|
廣島 |
西門答牛,褐色瑞士牛 |
|
|
|
短角牛,埃爾夏牛 |
|
|
取鳥 |
褐色瑞士牛,短角牛 |
|
|
島根 |
德盆牛,褐色瑞士牛 |
|
|
|
西門答牛,埃爾夏牛 |
|
|
山口 |
德盆牛,埃爾夏牛,褐色瑞士牛 |
|
|
愛媛 |
短角牛 |
|
|
大阪 |
褐色瑞士牛,西門答牛 |
|
|
鹿兒島 |
褐色瑞士牛,德盆牛,荷蘭牛 |
|
褐色和牛 |
高知 |
西門答牛,韓國牛 |
|
|
熊本 |
西門答牛,韓國牛 |
|
|
|
德盆牛 |
|
無角和牛 |
山口 |
亞伯丁安格士牛 |
|
短角和牛 |
青森 |
短角牛 |
|
|
岩手 |
短角牛 |
|
|
秋田 |
短角牛 |
由於各種外國品種被引進,並與各地區的本地牛進行雜交,而使本地牛的遺傳變異大量擴大。
到了 1950 年代中期以後,農業機械佔了優勢並且農業用化學肥料更為普遍而不需廐肥,因此役牛被取代而減少,這就產生促使飼養役牛轉向飼養肉牛。
現今黑色和牛在日本各地都可看得到。近來,在九州及北海道其頭數已在增加中;相對地在中國地區,曾經是主要生產黑色和牛的地區,目前其頭數則正在減少。
整體上該品種具有暗黑色的皮毛,有頭角而無肩峰,其身體大小有小型到中型的。成年母牛及公牛的身高與體重分別為
124 公分,420 公斤,與 137 公分,700 公斤。 180 天產乳量為 1,000
公斤。與其他的日本本地牛種作為比較,黑色和牛係以其牛肉產量特別著名,加上肉中帶有高度的大理石斑紋脂肪,且其皮下及內臟周圍的脂肪為薄薄的一層而已。
2.4 褐色和牛
(Japanese Brown) (如照片
5)

照片 5. 褐色和牛(公)。
褐色和牛品種有兩個截然不同的品系,一主要在熊本縣飼養,另一是在高知縣飼養。該兩品種的育成過程完全不同,所以通常分開敘述。
a) 熊本 (Kumamoto) 品系
原先在熊本縣有一種紅毛牛品系被飼養。該品系是從韓國進口的牛發展而成立的。在
1900 年之後,許多外國品種被引進,例如西門答牛和德盆牛,並與該品系雜交。該品系與西門答牛進行雜交,尤其會造出大型體格的牛。該品系的特徵是高速的增重率與大塊的肋眼面積。成熟母牛與公牛的體重分別為
600 公斤與 950 公斤。
b) 高知 (Kochi) 品系
高知品系是西門答牛與從九州島引進的韓國牛雜交育成。就進行雜交的期間來說,基本上高知品系比熊本品系短些。因此可減少稀釋原有品種的特性,且可保留重要的品種差異性。該品系亦具有黃褐色的皮毛,惟比熊本品系更為淺色。該品系牛具有黑色的角、蹄、眼皮、鼻鏡、舌頭、尾尖毛簇和肛門者,被認為屬於原有韓國品種的典型牛隻而受到更高的評價。該品系的牛肉生產性能與熊本品系很相似。成年母牛與公牛的體重,分別為
600 公斤與 950 公斤。
2.5 無角和牛
(Japanese Poll) (如照片
6)

照片 6. 無角和牛(公)。
無角和牛自
1916 年業已育出,係由本地的牛種與英國進口的亞伯丁安格士公牛雜交後育成。在
1975 年,為了改良肉質,曾以黑色和牛種公牛再與該品系母牛進行雜交。現在純種無角和牛的頭數可能所剩無幾。無角和牛的毛皮是黑色惟無頭角。自
1986 年後無角和牛一直未做能力或後裔檢定。無角和牛的成年母牛與公牛的身高與體重分別為
122 公分, 450 公斤及 137公分, 800 公斤。
2.6 短角和牛
(Japanese Shorthorn) (如照片
7)

照片 7. 短角和牛(母)。
短角和牛是雜交所造成。自
1871 年開始以本州島嶼(東北地區)北方的本地牛與進口乳用短角牛之間進行雜交,並聲稱此品種可利用夏季粗放的在該地區山嶽地帶放牧且比其他品種更為適合。現在主要分佈地區為東北和北海道,皮毛顏色呈現深紅褐色,比棕色和牛的顏色還深色些。短角和牛看來比黑色和牛更優於產乳、粗料採食及生長率。成熟母牛及公牛的身高與體重分別為
128 公分, 500 公斤與 140 公分,800 公斤。
3.
日本的本地牛品種與族群的遺傳分析
3.1 日本牛種的遺傳變異性
黑色和牛、棕色和牛和短角和牛等三個品種的遺傳變異,從數種指標,即使用血型、血液蛋白質、乳蛋白質和微衛星型 DNA 多態性當作基因標記等所得估值,其水準幾乎和荷蘭牛不相上下。另一方面,見島牛顯現低度遺傳變異性,幾乎是其他品種的一半 (阿陪等, 1977 加藤, 2000) 。口之島牛也和見島牛一樣,經使用相同微衛星型 DNA基因座顯示低度遺傳變異 (加藤, 2002)。
在 mtDNA , 透過母方品系遺傳,根據日本黑牛 18 個突變,有 24 個單種 (haplotype) 被觀察到 (滿年等, 2000),儘管見島牛在 6 個母方品系僅發現到兩個單種 (大澤等, 2002)。
阿部等 (1977) 報導無角和牛比其他的和牛品種具有稍微低度的遺傳變異。然而該品種的族群大小近來大大地減少且其遺傳變異的水平也似乎減少到關鍵水準。
每一種品種和族群有關的 Ppoly,平均異基因合子性和平均有效對偶基因,列於表 2.。
表 2. 五種日本的本地牛品種和族群的遺傳變異性
|
|
P.Poly |
平均對偶基因數 |
平均有效對偶基因數 |
平均觀測異基因合子性 |
|
口之島牛 |
56.5
% |
1.78 |
1.48 |
0.242 |
|
標準差 |
|
±0.85 |
±0.59 |
±0.248 |
|
見島牛 |
52.2
% |
1.78 |
1.40 |
0.209 |
|
標準差 |
|
±0.90 |
±0.48 |
±0.236 |
|
黑色和牛 |
87.0
% |
4.09 |
2.28 |
0.446 |
|
標準差 |
|
±2.21 |
±1.09 |
±0.281 |
|
短角和牛 |
95.7
% |
4.13 |
2.51 |
0.516 |
|
標準差 |
|
±1.74 |
±1.17 |
±0.234 |
|
褐色和牛 |
95.7
% |
4.48 |
2.90 |
0.560 |
|
標準差 |
|
±2.11 |
±1.35 |
±0.267 |
|
荷蘭牛 |
87.0
% |
3.78 |
2.51 |
0.521 |
|
標準差 |
|
±2.07 |
±1.23 |
±0.260 |
從日本本地牛族群的這些指標數據可以明確看出低於其他三種和牛品種。
荷蘭乳牛、口之島牛和見島牛的平均有效對偶基因分別為
2.51、 1.48 及 1.40,而其平均異基因合子性分別為 0.521、 0.242
以及 0.209。由此顯然可看出日本的純種本地牛的遺傳變異是十分低的
(如表
2.)。
3.2 從遺傳的距離所估計的日本本地牛各品種間與各族群間的遺傳關係
在 3 頭和牛品種、 2 頭日本純種本地牛族群及荷蘭乳牛品種找到 23 個微衛星型基因座的 142 個對偶基因。在見島牛及口之島牛的族群所觀察的只有 58 個對偶基因,而 58 個對偶基因中的 45 個即與他組,三頭和牛品種和荷蘭乳牛等所共有, 12 個對偶基因只與和牛品種共有,此外只有一個對偶基因是見島牛族群特有的。所以這表示現有日本肉牛品種依然存在著許多從前日本本地牛族群的遺傳基因。
從血型、蛋白質和 DNA 多態性所估計的遺傳的距離,指出和牛與荷蘭牛之間的關係為密切。至於日本的本地牛,即見島牛或口之島牛與和牛或荷蘭牛之間則顯出相當大的距離,並且見島牛與口之島牛之間的遺傳距離大過於日本的本地牛與黑色和牛或褐色和牛之間的遺傳距離 (表 3.)。
表
3. 日本的本地牛品種和族群的遺傳距離
|
|
口之島牛 |
見島牛 |
黑色和牛 |
短角和牛 |
褐色和牛 |
荷蘭牛 |
|
口之島牛 |
* |
|
|
|
|
|
|
見島牛 |
0.349 |
* |
|
|
|
|
|
黑色和牛 |
0.249 |
0.225 |
* |
|
|
|
|
短角和牛 |
0.382 |
0.383 |
0.196 |
* |
|
|
|
褐色和牛 |
0.251 |
0.254 |
0.104 |
0.183 |
* |
|
|
荷蘭牛 |
0.411 |
0.323 |
0.184 |
0.181 |
0.178 |
* |
雖然屬於純種的日本本地牛共有的許多對偶基因仍然留在和牛的各個品種,可是日本的肉牛的各個品種與荷蘭牛品種之間的關係,由遺傳距離推定,是相當密切的。縱使和牛源自日本的本地牛,在遺傳上卻與起源的牛種產生很大的差異,這是由於品種育成早期與外來品種雜交所致。
日本的兩種本地牛族群間顯示的大間隔,其原因,據推想,有些部分是由於引進兩座島嶼後遺傳浮動趨向不同方向所致,同時指出過去日本牛種族群中有地理上分佈差異。據報告這兩種族群有另類表型差異,如前述兩族群的體毛顏色不同。見島牛的肉質與黑色和牛相似,肉中具有有名的大理石斑紋脂肪沉積,而口之島牛則產瘦肉而已。黑色和牛具有大理石斑紋肌肉似乎來自中國地區所飼養的本地牛,該地區就是黑色和牛的育成地,同時也是見島牛引進原地。
4. 日本的本地牛展望
日本育成四種肉牛分別為黑色和牛、褐色和牛、無角和牛和短角和牛。實際上還是把這些品種看做日本的本地牛,儘管在 1900 年初起初廣泛與外來牛雜交過。雜交所利用品種以及所使用選拔標準各縣顯然有異。結果育成一些特殊的品系。然而現在遺傳變異減小,這是由於人們集中在數目有限的黑色和牛品系上,注意具有上等肉質所致。 1991 年由於牛肉自由進口之後,另有其他品種,因其肉質被認為很難與外國肉牛分出優劣出來,其飼養頭數就急劇減少。
由此,在現階段日本的本地牛可分類為三類: 1. 還沒有危險的黑色和牛 2. 現在沒有危險但未來可能變成絕種的,褐色和牛與短角和牛 3. 在危機中,無角和牛、見島牛及口之島牛。從遺傳保種觀點看,要有系統的保存少數日本的品種和少數黑色和牛品系,(作者)建議,為了未來遺傳資源的需求上,使用冷凍精液及胚胎當作緊急方法或現存動物的保種補救方法。
從永續利用牛的品種及充分利用日本自然資源而沒有環境損害等的觀點來看,發展適當的飼養方式以供上述各種牛品種及兩種本地牛種群是必要的。在熊本縣褐色和牛和東北地區的短角和牛,具有優良食草性狀,與黑色和牛相比,是在多山丘陵地區育成並放牧長大的。在
2000 年,直接支付系統已在山丘地區依照食物農業與農村基本法開始施行。在阿蘇,該項補助款得使用於提昇動物生產能力,集中在維持和管理牧草地及褐色和牛品種的工作上。
參考文獻
Many parts of descriptions are based on Obata et al. (1996).
Obata T, Satoh M and Takeda H (1996) Conservation and Utilization of Beef Cattle Genetic Resources in Japan. Animal Genetic Resources Information 18: 65-78.
Abe T., Oishi T. and Komatsu M. 1977 Genetical Constitution of Japanese cattle as determined by the Gene Frequencies of Blood Groups and Protein Types. (In Japanese with English summary) Bulletin of National Institute of Animal Industry 32: 63-69.
Furukawa T., Nirasawa K., Izaike Y.and Noguchi J. 1997 Investigation of Animal Genetic Resources in Yamaguchi Prefecture 1. Investigation on conservation of Mishima Cattle in Yamaguchi Prefecture. (in Japanese with English summary) Survey Report for Animal Genetic Resources 7:51-76.
Harada, Y., Sakata, S., Akimoto, I., Fukuhara, K., Shinoda, T., Hosoi, E., Ozawa, S. (1996) Experiment for the performance of Mishima Cattle. Proc. 8th AAAP Anim. Sci. Cong., 2: 356-357.
Hayashida and Nozawa (1964) Cattle of Tokara Islands. (in Japanese) Report of the research group on the native farm animals in Japan and its adjacent localities 1:24-29.
Japan Livestock Industry Association (2001) Statistical data related to livestock improvement. Livestock Improvement Association Japan, Tokyo, pp. 155.
Kato K 2002 Studies on Genetic Variability of Japanese Native Cattle based on Microsatellite DNA Polymorphism. (in Japanese) Master’s Thesis, Graduate School of Bioagricultural Sciences, Nagoya University, pp 72.
Mannen H, Kawasaki J, Ishida T, Mukai F and Tsuji S. 2000 Mitochondrial DNA Diversity of Japanese Black cattle. (in Japanese with English summary) Animal Science Journal 2000 71:J470-J474
Nishida S (1983) History of Animal production in Japan, “Treatise on Animal Production –Pantography of Animal production” (in Japanese), Rural Culture Association, Tokyo, pp. 125-267
Shi J, Hosoi E, Harada Y, Miyake S, Sakata S, Shinoda T and OZAWA S 2002 Genetic Variants of Mishima Cattle Based on Analysis of Mitochondrial DNA D-loop Region (in Japanese with English Summary) Nihon Chikusan Gakkaiho, 73:261-264
Tomita, 1996 Japanese Feral Cattle (in Japanese) Chikusan No Kenkyu 50: 125-129.
Tsuda T (2001) Cattle and Japanese People – An Attempt of Cultural History of Cattle (in Japanese). Tohoku University Press, Sendai pp300.