��F�|�A�~�e���|�b������ҤE�Q���~�Ĥ@�u�dzN��Q�|(2006/03/14)
��]���X�ƪ��ʪ��|����t
DNA Barcode System for Genomic Breeding Animal
�d�����B��౹šB�C���O�B�����_
��F�|�A�~�e���|�b������ҿ�Ǩ|�ز�
mcwu@mail.tlri.gov.tw
http://www.angrin.tlri.gov.tw
2006/03/14
�@�B�~���v�O�@���{���αM�Q�O�@��]��
�G�B�b���~�ؤΫ~�t���Q��
�T�B��V����B�ʬV����βɽu���]
�|�B�دb�V�Ͳ��i���ΰ�]�n��
���B��]��ؤα��X�ƪ�����
���ΰ�]��اN���|���~�ؤΫ~�t�A�ŦX�u�~�ءB�~��B�~�P�v�@�馨�����W�~�A�~�A�]�i�i��U�~�A�~�u�~�ޡB�~���B�~�y�v�������Q�ΡC�ʪ��|����t�]�A�X�͵n�O�B�ʯ��˩w�B�إεn���κدb����Ų���|�j�D�b�C��]���X�]�A��V����B�ʬV����βɽu�鵥��]���ͪ��Ӽе��U�A�ϥΰ�]�ǦC���t���ӽs�X�A�D�n�O��ǤW����@�P�����ίS�w�P��ǦC�����ơC��]�O�ͩR�骺���[�A�}�i���ҬO�O�ٰ�]�\��A�{�A�ӤH����|����J�O�l�D�~��í�w�פΥͲ��įq���@�C
�@�B�~���v�O�@���{���αM�Q�O�@��]��
�~��(Breed)�P�~�t(Line��Type)�����O�A�~�ػݭn���㪺�~���S�x�ӰϤ��A���~�t���Ϥ��h�����q�~���W�ӧP�O�C�~�ض��㤣�P���~���S�x�A�~�t����ۦP���~���S�x�C�~�ؤ����ʯण�P���~�t�C�۵M���ҩҭl�ͪ��ڸs�A�ڭ̷|�٨䬰����(Species)�ΨȺ�(Subspecies)�A�ӫ~�ػP�~�t�O�g�L�H���ѻP���B�J�ӧΦ����ڸs�C�b���k(���إ���87�~6��24���`��(87)���`(�@)�q�r��8700123980���O��w�����B���إ���94�~1��19���`�ε��`�@�q�r��09400004871���O�ץ����G)�ĤQ�G���u�o�{�B�|���Φ۰�~�i�s�~�ةηs�~�t���دb�V�κح�̡A���V�����D�����ӽеn�O�A�g�f�w�֭�n�O��A�l�o���s�B�P��C�v�C�b���k�ĤG�Q���u�����D�������O�s�دb�V�귽�Χ�}�a�b�B�a�V�ʯ�A�o�e�оdzN��s���c�Υ�������q�Ʀ����BŲ�w�B�O�s�ά�s���ƶ��C�v�C�b���k�ĤG�Q�@���u�����D�����o�w����Ų�دb�V�~�̡C�g��Ų�u�}�̡A�������y�C�v�C
��F�|�A�~�e���|���ߴӹA�����߳��A�N��a��J�������o���G�L�v���ѹA���ϥΡA�����]�ʥF�M�Q�O�@�A�y���ꤺ�A�~��}��o��ެy���A�����ζ�������x�W�A�~�g�ٳy���¯١C�g�ٳ����z�]���������m�ͩ�2006�~1��19����ܡA�����ɥx�W�A�~���[���ȩM����v���O�A�g�ٳ��ۤ�q�u�M�Q�k�v�A�N�}��ʴӪ��M�Q�O�@�A���y�ʴӪ��~�ج�o��}�C�ثe�H����B�饻�B�s�[�Y���ʴӪ��M�Q�O�@�̰����A�a�ʰ�]��ެ�o�i�F�ڬw��a�ȭ���}��Ӫ��M�Q�O�@�C����ڤj�ͶդU�A�V�ӶV�h��a�M�w�}��ʴӪ��M�Q�O�@�A�x�W�����[�t�}��C
�ʴӪ��M�Q�F���W�A��F�|����U�ݲթ�2005�~8��23��o�G�s�D�Z�A���D���u�F�|�M�}��ʴӪ��M�Q�O�@�A�ɥͧP�A�~�¥��~�Ƶo�i�v�C�P�~12��6��ͪ�IPR(���z�]���v)�u�@�p�ղĤT���e���|ij���ʴӪ��M�Q���סu�ʴӪ��M�Q�F���w���O�ѹA�e�|�Φ�F�|�ͪ��N���~���ɤp�սT�{�A���z����̾ڦ������ʴӪ��}��M�Q�������תk��סA����д��z���t�X�M�Q�k�תk�i�i�椽ť�|�Ϋ���ߪk�u�@�C���йA�e�|�t�X�]���ʴӪ��}��M�Q���������q�Ϋžɤu�@�v�C�]���A�g�ٳ����z�]����������ժ���2006�~1��24��b�A�e�|�����|�W�A�����}��ʴӪ��M�Q�����M�Q�k�ץ������סC�w��ʴӪ��M�Q�n�O�@����H²��Ө��A�~���v�O�@���{���A�M�Q�O�@��]���C�ҥH�����ק�M�Q�k����24���B��57���β�78���N�i�H�}��b�x�W�ӽаʴӪ��M�Q�C���ӡu�M�Q�k�v�}��ʡB�Ӫ��ͪ��N�o���M�Q�A���p�i�W�[��d�������A�~���ȡC���z�]�����N�M�Q�k����24���B��57���β�78���u�M�Q�k�ץ���סv�e��F�|�f�d�C�ץ����e�]�A�ʴӪ��M�Q�}�d��B�|�خa�K�d�v�B�v�Q�ӺɡB�A���K�d�v�B�M�Q�v���j����v�Υ椬���v���C
�M�Q�k����24���O�W�d�ʴӪ��M�Q�}�d��A�ץ���סG�u�U�C�U�ڡA�����o���M�Q�G�@�B�H��ΰʪ��e�f���E�_�B�v���Υ~���N��k�C�G�B���`���@���ǡB���}���U�νå̡ͪC�v
�M�Q�k����57���O�B�z�ʴӪ��M�Q�|�خa�K�d�v�B�v�Q�ӺɡB�H�ιA���K�d�v�A�|�خa�K�d�v�ץ���סG�u�o���M�Q�v���ĤO�A���Ω�U�C�U�ڱ��ơG�@�B�ӤH�D��Q�ت����欰�C�G�B����s�B�����I��o���C�T�B(��)�v�F�W�q��57����1�v�Q�Ӻɯ�סG�u�M�Q�v�H�һs�y�θg��P�N�s�y���ͪ����Ƴc���A��M�Q�v�ĤO���Ω�ӥͪ����Ƹg�c�ީμW�ަӪ�����o���ͪ����Ƥ��o���c�ީμW�ޥت��A�ϥΡC�e���ͪ����ơA���H�c��ᥲ�M�ɭP�ͪ����ƪ��c�ީμW�ު̬����C�v�F�W�q��57����2�A���K�d�v��סG�u�M�Q�v�H�һs�y�θg��P�N�s�y���M�Q�Ӫ��c�ާ��Ƴc���A��M�Q�v�ĤO���Ω�A�����c�ީμW�ޥت��d�ئۥΤ��欰�C�e���Һ٤��Ӫ��H�Ӫ��~�ؤκح]�k��26�����i���Ӫ����ج����C�M�Q�v�H�һs�y�θg��P�N�s�y���M�Q�ʪ��ΰʪ��ʹާ��Ƴc���A��M�Q�v�ĤO���Ω�A�����A�~�ت��ϥθӰʪ��ΰʪ��ʹާ��ơC�����]�A���ӷ~�ʹެ��ʥت����c��欰�C�v
�M�Q�k����78���O�W�w�M�Q�v���j����v�Υ椬���v�A�W�q�ڶ���סG�u�~���v�H�Q�Ϋ~���v������I�L�H���ͪ��N�M�Q�̡A�o�̲ĤC�Q�����W�w�ӽЯS�\��I�C���ӫ~�ض��M�Q��۷��g�ٷN�q�����n�N��}�̡A�l�o�ӽЯS�\��I�C�M�Q�v�H�̴Ӫ��~�ؤκح]�k���o�~���v���S�\��I�~���v�̡A�~���v�H�o�P�M�Q�v�H��ij�椬���v��I�C�~���v�H���o���S�\��I�v�A���P��~���v�@�������B�~�ӡB���v�γ]�w���v�C�v
�G�B�b���~�ؤΫ~�t���Q��
�ͪ���ǯS�ʫY�۵M�ɪ��ت�������Ϯ����Ҥ��U���ͦs�D�ԻP�O��v���Φ��A��Ǫ��h�˩ʬO���ئs�n����A��O�H���i��~�ا�}�P��|���D�n�ӷ��C��~�ӥ@�ɦU��}�l�������@�A�~�@���B�b�V�B���Ͱʪ�����Ǹ귽�h�˩ʡA�H���ѩ�L�����g�٧@���P�b�V�B�����ͪ����g�٩ʯध��|�A�\�h�~�ةΩʪ��]�ӾD�쩿���Ӯ����A���H�Y�ǫ~�ؼ�b����Ȥ���Ǹ귽���_�s�b�A�]�ӳy���L�q�������l���C�ڰ���O�H�A�~��ǡB�Ӫ����`�B�f��ͪ��B�L�~��ǡB������ǡB�b�V��ǡB�~��f��L�ͪ��P���Ͱʪ���ǵ��K�j�ح�w�A���[�j�ح줧�S�ʽլd�B�����θ�ƹq���ơA�H�����ح���ƧQ�ΡA�F��c�اڰ꧹�㤧��a�A�~�ح�O�s�Q����t�A�T�O�A�~�������o�i�C
�DzΨ|�ج��ڰ�A�~��o�κدb�V���~�o�i���u�թҦb�A�i�@�B�}��ʪ��M�Q�v�ӽСA��b���ح�O�ءB�|�ؤΰ�]��صo�i���G���X�O�١A�åi�P�i���~�ɯŤΪ����૬�A���Q��p�ڸs�|�ؤΦh�˦�����զX�s�~�ةηs�~�t�C
�b���k�ĤQ�G�����@�u�دb�V�κح�A�ο�Ǫ�����m�̡A�������ж�����Υͪ��w���ʵ����A�l�o���s�Q�ΡF���Ǫ�����m���z��k�A�Ѥ����D�����w���C�v�C���إ���91�~11��15���F�|�A�~�e���|�A���r��0910040491���O�q�w�o���u��]��دb�V�ж�����Υͪ��w���ʵ����z��k�v�C���إ���94�~1��26���F�|�A�~�e���|�A���r��0940040111���O�q�w�u��]��دb�V�κط���X�J�P�N���f�֭n�I�v�C
�b���k�ĤQ�G�����G�u�G�H�H�W��P�@�s�~�ةηs�~�t���دb�V�κح�H�ۦP�Ϊ�����W�٦U�O�ӽеn�O�ɡA����̥��ӽЪ̵n�O�F��b�P��ӽЦӤ����O����̡A�ѦU�ӽФH��ij���k�@�H�M�ΡF����F����ij�ɡA�H���Ҥ覡�M�w���C�G�H�H�W�دb�V�~�̩k�I��e�w���s�B�P�⤧�~�ةΫ~�t�A�H�ۦP�Ϊ�����W�٦U�O�ӽеn�O�ɡA�����D����������|���εo�{�g�L�ι}�i���i��O����M�w���C�����H�Ҩ|���εo�{���s�~�ةηs�~�t���دb�V�κح�A�������t�����w�~�A���H���ΤH�W�q�n�O�C�v
�b���k�ĤQ�T���u�̫e���n�O���~�ةΫ~�t�A�g�����D�������w���i�̡A�دb�V�~�����V�����D�������w�����c��z��εn���C�v
�b���k�ĤQ�E���u�g�����D�������i���دb�V�B�ط��A�����o�����D�������P�N���A�l�o��X�ο�J�C�v�C���إ���92�~2��14���F�|�A�~�e���|�A���r��0920040147���O�q�w�o���u�دb�V�κح��X�P�N���f�֭n�I�v�A���إ���94�~4��15���F�|�A�~�e���|�A���r��0940040251���O�ץ��u�دb�V�κح��J�P�N���f�֭n�I�v�C
��1987�~�}�l�i��ڰ�b���ح�O�s�ΧQ�ΡA��覡��������Y��O�W�b����ƩM�A����Ƥ��t�ܡA�䭫�n�ʤ��Ȧb�O�@�P�c�O�W�S�������a�ةκ٦a���(landrace)�a�b�V�ڸs�A�O�����ޡB�����ޡB�����B�����B�¤s�ϡB�g���B�Ŧ���n�B�¦�f�n�B�g�Z�B������M�������x�W�S�����~�ءA�åB�O�s�F�S�w�S�ʿ�|�������ީM�զ���n�s�~�ءC�ح�O�s�ΧQ�άy�{²�C��U�G
(1)�ح�~�إ~���S�x�зǤ��q�w�C
(2)�ح�˥����ʡC
(3)�c���Ҥι}�i�覡����w�C
(4)�O�رڸs�ij{���t�ؤ覡�c�l�U�@�N�C�Y�w��Y�@�S�ʤ��¤ơA�h�i��¨t��|�ڸs���إߡC�Y���}�o�ݨ㥻�a�M�i�f�~�دS�ʤ��s�~�ءA�h�إߥt�@�S�ʱڸs�ѯŶi�|�ة������|�C
(5)�����Ƥ������A�]�A�髬�ܤơB�o�|�����L�{�B�c�ު��p�Φ欰�ߩ��[��C
(6)���ΥͤƾǩM�ӭM�ǧN���R�O�دb�V����G�Ͳz�ȩM�V����֫��C
(7)�Q�ΧN��O�s�N�ӫO�s��l�B�Z�M�F���ʹӭM�C
(8)�إ߭��鶡�t�и�ƤΫO�s��Ǫ���A�Ѱl�a��ǯS�ʥΡC
(9)�s�L�ح�O�s�ΧQ�Τ�U���媩�έ^�媩�B��p���ɡB�O�ؤ��L��B�a�b�V�ʺA�ͬ��v���v�a����s�H���c���Ҥ����[���u�@�A�H�P�Ϥj���糧�g�b���귽���i�@�B���{�ѡC
(10)���ڸs�ƥ��X�j��ίS�ʿ�|�ᤧ�~�عF�@�w���ǮɡA�h���ѵ������~�̶i��j�q�c�ޥͲ��C���ɡA��i�z�L�ح�洫�D�A�i���ڶ���y�C
|
���@ �b���ح�w���~�ءB�ƥػP���a(1987~1992) |
||||
|
�ح� |
�~�� |
�O�رڸs�ƥ� |
�O�س��a |
|
|
�� |
���� |
10�� |
30�� |
�b�թ��`�� |
|
������ |
15�� |
45�� |
�b�թҥx�F�� |
|
|
������ |
6�� |
21�� |
�x�j�b���Ǩt |
|
|
�� |
���� |
2�� |
30�� |
�b�թҪὬ�� |
|
���� |
11�� |
70�� |
�b�թҫ��K���� |
|
|
�� |
�¤s�� |
5�� |
50�� |
�b�թҫ��K���� |
|
�¤s�� |
4�� |
40�� |
�b�թҪὬ�� |
|
|
�� |
�g�� |
200�� |
800�� |
�b�թ��`�� |
|
�g�� |
35�� |
100�� |
���j�b���Ǩt |
|
|
�n |
�Ŧ���n |
80�� |
200�� |
�b�թҩy������ |
|
�զ���n |
150�� |
450�� |
�b�թҩy������ |
|
|
�¦�f�n |
70�� |
70�� |
�b�թҩy������ |
|
|
�Z |
�Ŧ�g�Z |
40�� |
120�� |
�b�թҹ��Ƴ� |
|
�զ�g�Z |
40�� |
120�� |
�b�թҹ��Ƴ� |
|
|
�� |
����� |
16�� |
25�� |
�b�թҰ����� |
|
���� |
9�� |
13�� |
�b�թҰ����� |
|
��1998�~�q�w��I���b���k�ĤG�Q���u�����D�������O�s�دb�V�귽�Χ�}�a�b�B�a�V�ʯ�A�o�e�оdzN��s���c�Υ�������q�Ʀ����BŲ�w�B�O�s�ά�s���ƶ��C�v�A�����ح�~�责�@�Φh�˩ʦ��䥲�n�ʡC�G��b���ح�p�e�b2007�~����20�~���e�A�|�ݥ[�j�Q�Υ��a�ؤ��g�ٯS�ʿ�ޡA�����a�~�ةM�~�ӫ~�ؤ��S�ʿĦX���s�~�ءC�ӫO�س����H�p�ڸs�c�ޤ覡�Өt�C�ʰO���~�[�S�x�B�ͬ��ߩʡB�g�٩ʪ��B�Ͳz��ǭȩM�����|�L�{�~�A�öi��ʹӭM�N��O�s�u�@�M�ʪ��^�y�����_�|�u�@�C
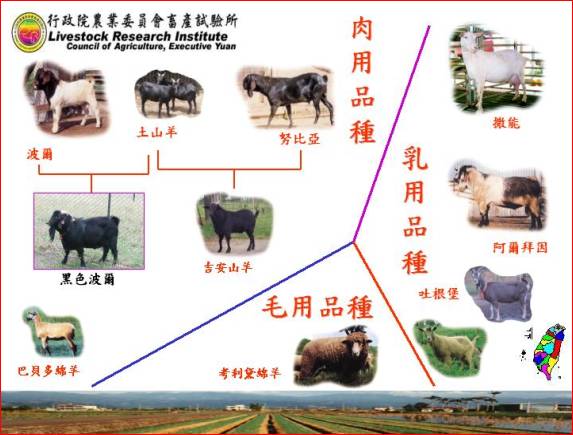
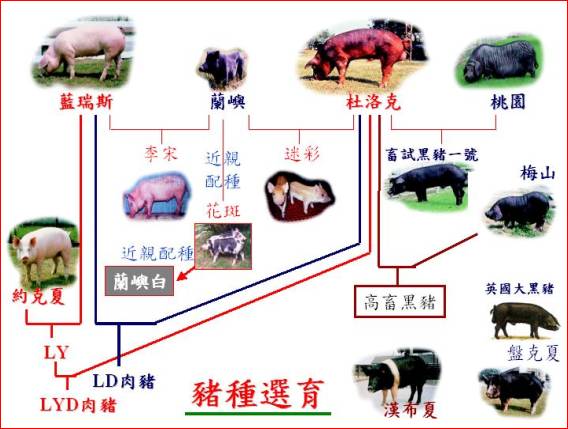
�s�~�سq�`�O�ѥ~���S�x���P����~�ءA�ƦܤT�ӫ~�إH�W������ҥͫ�N�A�A�̾ګ�N�~���M�ʯ�i��D�אּ�إο˥N�A�ۦ��c�ް��|�X�s�@�N���ڸs�A�㦳�N�N�۶Ǫ��~���A��~�������i��Ȥ@���~���S�x���P���˥N�A�h�i�����s�~�ءC���b1975�~�x�W�j�ǯb���Ǩt���n���б¤Χ��øq�б����a��ǩe���|��s�p�e�ɧU�A�Q�Φa��~���������ްt�~�Ӻ��ŷ紵�ު���~������A��1983�~��~�žɤw�|���զ�p���ޡu�����ޡv�F�A�p�Q�Φa��~�خ��P�~�Ӻا����J�ު���~������A���طs�X������N�g�ۥ��|���t�@�W�߱ڸs�~�ءu�b�ն½ޤ@���v�F�H�ίb������ҥx�F���Q�Φa��~�������P�~�Ӻا����J�ު���~������A�H��⦳�Ħ������M�զ������۶������P�M�S�f�t�A�|���p���ު�˨t�ư���0.60�s�~�ءu�b�հg�m�ޡv�C
�t�@�ب|���s�~�ؤ�k�O�q�a��رڸs����ޥX�ܲ�����A�q�`�O��⪺�t���A�Ҧp�b������ҩy�����Ҧ۽Ŧ���n�s�D�X���馳�dz\�զ�Ф���A�v���令95%����⬰�զ�u�y�����n�x�b�@���Υզ���n�v�C�b������ҥx�F���]�q�¦������ޫO�رڸs���D�X���馳�dz\�զⴳ������A���Υb�P�M�t�ؤT�N��A�A�Υ��P�M�S�f�t�令�զⴳ���������B��˨t�ư���0.60�s�~�ءu�b�ժᴳ�ޡv�C
�s�~�t�x���a���(landrace)���ʪ��b�Y�@�a�ϸg���ӥ@�N�A���צ��L�H����ޡA��O�رڸs�w�A�����a��Ԥι}�i�z�覡���~�A�٥i�H�c�ޫ�N�h�~�A�Χe�ʳ��ڸs��15�~�H�W�A�w�ۦ��@�өʯ�í�w���ڸs�A�]���i�����s�~�t�A�Ӥ��H�R�W����s�ܥL�a�C�]���A�Ѧp�b������ҪὬ�����x�W�����ίb��������ڬK���Ҫ��x�W�����A�i�Q�����a��ت��s�~�t�C�o�ئa��ظg�ʯ�O���ι}�i�c�ަ��p�ڸs�A�q�`�H�}�i�a�W�ӽЩR�W���s�~�t�C�b������Ҧۥx�W�U�a�����g���A�A���Υ��P�M�t�ؤ�k�Υ~���ʯ��ަӨ|���x�W�g����˫~�t7�B9�B11�B12���|�ӡC�A�p�b������ҩy�����Ҩ̲��J�ʯ��ޥX�u�Ŧ���n�b�դ@���v�ίb��������`�Ҧ۽Ŧ���n�b�դ@���ڸs��|�X�������ʰ����u�Ŧ���n�b�դG���v�C�ڰ�۰�~�ءA���O���A�����ҫ᪺�ʯध�~�A�٥i�H�A��|�X�A���x�W��Ԫ��s�~�t�ίS�w�ʯ�¤ƪ��s�~�t�C�θg�ѦP�@�~�ت���ӫ~�t�����A�]��|���t�@�ʯ�í�w���s�~�t�C
|
���G �O�W�b�V�ʪ��ح�(2006.03.01�s��) |
||
|
���a�� |
�b |
�O�W�����B�O�W�����B�O�W�¤s�ϡB�O�W�Ŧ�s�ϡB�O�W������B�O�W�����B���ޡB������ |
|
�V |
�o�ܤg���B�Ὤ�g���B�˱T�g���B�����g���B�����g���B�H�q�g���B�Ŧ�g�Z�B�զ�g�Z�B�Ŧ���n�B�¦�f�n |
|
|
�~�Ӻ� |
�b |
�����Ť��B�t�F���B���Ҥ��B�L���Q���B�w�洵���B�խ����B�V��Ȥs�ϡB����s�ϡB�������]�s�ϡB�R�ڳ��s�ϡB�i���s�ϡB�ҧQ�L�����ϡB�ڨ��h���ϡB�º����p�����B�k�Ԩ��g�A���B�æ����ըߡB�p�J���ߡB�ŷ紵�ޡB���J�L�ޡB�����J�ޡB�~���L�ޡB�L�J�L�ޡB���s�� |
|
�V |
Ī�����B�ú~�����B�Ӧ����B�L�����B���ЯQ�����B�_���n�B�զ�f�n�B�����B�m���B��ù���Z�B�g�|���Z�B�D�w�¤��Z�B�פ��Z |
|
|
�s�~�ث~�t |
�b |
�����ޡB�b�ն½ޤ@���B�b�հg�m�ޡB�b�ժᴳ�� |
|
�V |
�y�����n�x�b�@���B�Ŧ���n�b�դ@���B�Ŧ���n�b�դG���B��ˤg���~�t�C�B��ˤg���~�t�E�B��ˤg���~�t�Q�@�B��ˤg���~�t�Q�G |
|


��F�|�A�~�e���|�b������ҥ����|���s�~�ؤηs�~�t��10���p�e�A���Ǥw�������a�ж�����A���ǥ��w�ƿ�ǫ�í�w���˴��A�]����n�}�l����حp�e�C�s�~�ئW�٤ηs�~�t�W�ٷ|�̿�إؼйF���צӳv���ק�ܲŦX�Ͳ��t�ΩһݡA�G���X�R�W�e�᪺�W�ٷ|���h�ӧ��ܡC
|
���T ��F�|�A�~�e���|�b������ҥ����|���s�~�ؤηs�~�t |
||
|
�����|���s�~�ؤηs�~�t |
2006�~�p�e�D���H |
���|���� |
|
(�@)�s�~�t�|�� |
||
|
�զ�f�n�b�դ@������| |
�B�q�w�D�� |
�b�թҩy������ |
|
�b�ե�ù�Ҥ@���ΤG������| |
�d��ܥD�� |
�b�թҹ��Ƴ� |
|
�b�դg�������յg�~�t����| |
��q�K�U�z��s�� |
�b�թҿ�Ǩ|�ز� |
|
�x�W�����Ὤ�~�t����| |
�Q�}��U�z��s�� |
�b�թҪὬ�� |
|
�x�W�������K�~�t����| |
�����_�U�z��s�� |
�b�թҫ��K���� |
|
�h�������J�ޫ~�t����| |
�d�����ժ� |
�b�թҿ�Ǩ|�ز� |
|
(�G)�s�~�ب|�� |
||
|
���b�½ޤ���| |
�\�ʻ��D�� |
�b�թҰ����� |
|
�զ������ޤ���| |
�����y�D�� |
�b�թҥx�F�� |
|
�b�զN�w�s�Ϥ���| |
���z�اU�z��s�� |
�b�թҪὬ�� |
|
�b�ն¦�i���s�Ϥ���| |
Ĭ�w��Ƭ�s�� |
�b�թҫ��K���� |
��ڹA³�Ӫ��ح��w(International Treaty on Plant Genetic Resources for Food and Agriculture)�A��2004�~6��29�饿���ͮġCñ����N���q�ȫO�s�@��³�A�@�����h�˩ʤμi���w�����e�C�ھڬ����έp��ܡA�q�H�����A�~�H�ӡA�ܤ֦�10,000�إH�W���Ӫ����ش��Q���@�����έ����Ӯ�ءC����A��150��³���@���ѩҦ��H��³�����ӷ��C�ӳo150��³���@������12���A�S���ѤF�H��80%�������ӷ��A�b�o80%���A�p���B�_�̡B�ɦ̤ΰ��a���A�S���F60%�C�Ѧ��i��³�A�@�����ͪ��h�˩ʥ���@��ֻP�y���C�]���A���غح쪺�O�s����ݫإߡA�ӥB�u���z�L�s�x���Q�ΦU�ت��ت��ح�A�~��T�O³�A�@���h�˩ʪ��}�o�A�H��i�H��³�������O�C�p�X��³���ιA�~��´(FAO)��2000�~�a�b�V�~�إؿ��A�Ȭw�Τӥ��v�a��1999�~���a�b�V�~�ئ����@�ɪ��ʤ���H�����B�ޤ��n���F30%�H�W�A��ܳo�a�Ϧ]���U�Φa�z���Ϲj�A�ɭP�p�ڸs���c�ޤ覡�~�h�C��~�ӡA��ڶT���ۥѤƤβ{�N�}�i�N���Ǽ����ΡA�ϱo���ǫ~�ئ]����g�ٻ��ȡA�Ӥ��A�c�l�A�Ѧp���@�ɪ��s�ϫ~�ثܦh�A�h�F587�ӫ~�ءA�䤤��17�ӫ~�ؤw���ءC���Ϫ��~�ا�h�F1495�ӫ~�ءA�䤤��181�ӫ~�ؤw���ءC
|
���| �Ȭw�Τӥ��v�a��1999�~���a�b�V�ƶq�B�~�ؤΦ����@�ɪ��ʤ��� |
||||
|
�ʪ��O |
�Y��(�Y) |
�~�ؼ� |
�����@���Y�ƪ��ʤ��� |
�����@�ɫ~�ؼƪ��ʤ��� |
|
���� |
152,404,000 |
61 |
93 |
70 |
|
�� |
461,197,000 |
236 |
35 |
19 |
|
�s�� |
390,433,000 |
146 |
55 |
26 |
|
���� |
408,098,000 |
233 |
39 |
18 |
|
�� |
525,598,000 |
184 |
55 |
37 |
|
�� |
14,859,000 |
83 |
25 |
11 |
|
�j |
14,885,000 |
12 |
34 |
12 |
|
�d�m |
2,815,000 |
14 |
15 |
22 |
|
�� |
6,181,645,000 |
124 |
45 |
18 |
|
�n�εf�n |
717,811,000 |
45 |
92 |
45 |
|
�Z |
189,436,000 |
13 |
90 |
20 |
|
���� |
2,142,000 |
6 |
1 |
18 |
|
��ƨӷ�: FAOSTAT (estimates of 1999 live animal populations) and DAD-IS |
||||
�x�W���Ť��B�ީM�J���~�ؤ��巽���ڬ��餧�ӥΫ~�غح�b���A��~�����s�巽�i�A�g�L�ƤQ�~���x�W��Ԥ��۵M�z��A�w�t���A���������Ԫ���]�A�[�W�h�~�ӥx�W�����o�ǫ~�ت��g�ٻ��ȤΥͲ��IJv���@�A���x�W�b�V�~�ؤw�������a�a�Ϫ��u��~�t�C
�T�B��V����B�ʬV����βɽu���]
�ʪ�����Ǫ���s�b����V����B�ʬV����βɽu��A�H�h��ֿ}�ֻ�(DNA)�覡�s�b�A�b�ӭM�֤�����V����DNA�ΩʬV����DNA���u���A�Ӧb�ӭM�誺�ɽu�餺DNA�h�������CDNA�O�Ѯ֥̻Ī����骫(�U���P��A�^��Base pair, bp)�Ҳզ��A�C�@�Ӯ֥̻ħ����C�ġB���ҿ}�Υ|�Ӵ��Q���@�ӲզX�Ӧ��C�|�Ӵ��Q�A���I��(A)�M���T�I��(G)�G�̬O�ݩ��I��ڡF�ݸ��G�r(T)�M�M�G�r(C)�G�̫h�ݩ��G�r�ڡCDNA���Ѹ̡AA�MT�H��ӲB��ۧl�A��G�MC�H�T�ӲB��ۧl�C�bDNA����ѧǦC���A�t�s�X�ϰ�(�q��Exon)�P���s�X�ϰ�(�q��Intron)�C�s�X�ϰ쪺DNA�ǦC��Q��Ķ���J�ս�A�o�dzJ�ս�㦳�Ͳz�\��F���s�X�ϰ쪺DNA�ǦC�h�L�k��Ķ�X�J�ս�A��ͪ��\��|�������M���A�M���ϰ�t���j�q������ܲ��סA���i�ΥH�ϧO���鶡����Ǯt���ʡC
DNA���N(�U�ٮֻ��˴��ΰ�]�˴�)���\�h��k�A�p�����k(Fingerprinting)�B����ʤ��q���צh�A���k(Restriction fragment length polymorphism, RFLP)�B�L�ìP���w���k(Microsatellite typing)�B�g�A�ìP���w���k(Minisatellite typing)�M��Ǧ줸���R�k(Genetic bit analysis)�A�C�Ӥ�k�U����d��P�ɪŭI���C�L�ìP�O���bDNA�ǦC���A�t���u�����p���ƪ��֥̻ġA�䭫�ƪ���즳��B���B�T�Υ|�Ӯ֥̻ġA�Ҧp���O��TTTTTT�BTGTGTGTG�BGTCGTCGTC��GATCGATCGATC�b�o�DzզX���A�H�����ƪ̳̦h���C�o�Ǧ��p���ƪ��֥̻ħǦC�q�`��s�X�ϰ�A���ϰ�t���j�q������ܲ��סC�L�ìP�w���k�����R�A�]�t�����D�`�M�~��DNA���q�M�p��Ӱϰ줤���ﰸ��]�j�p��譱�C���ؤ�k�����I�b���o�ǷL�p���t���ɡA�ݭn�۷��Y�V������N�F���u�I�b��ϥΦ��k����e�{�ʪ������j������ܲ��סA��۷��e���h�����M���ΡC���ϥλE�X�Q�s�����(PCR)�ӧֳt�i��DNA���ɡA�L�ìP���ޤl(Primer)���]�p�S�O���n�A�����N�ޤl�]�p�b�u�B�S�w��DNA���q���d�A�åi���T�a����ﰸ��]���j�p�C����PCR�ӿz�ˤ��P���骺DNA�A��N�ܤֶq��DNA�g�L���Ъ��@�Ϋ�A��ϯS�wDNA���q�ƻs�W�q�A�A�H����q�a�����V���e���οå��۰ʤ��R�����A�o�H�P�_��j�p�ΧǦC�t���A��i�N��DNA��ﵲ�G��ͪ��c�ީʯ��ơA����]�y�s�ꪺ���R�A��X�g�٩ʪ���]�C���Ҧ����Q��]�[�H�զX���@��Ʀ�X�A�N�O�ӫ~�ث~�t����]���N�X�A��٬��u��]���X�v�C
��]���X�һݪ�DNA�s�X�N�����ΦU���ؤ����S�w�H���A�Ʀܩ���鶡�S�w�H���ADNA�W�s�b�۳\�h�����苷�Ħh�A�� (Single nucleus polymorphism, SNP) �զX�A�Φ����νs�X���ʲӤ��P���u���C�ʪ��ɽu��DNA���t�Ƴt�v�۹��V���骺DNA�֫ܦh�A�]���Q�Ψ�Ų�w���ءA�l�N���ɽu��DNA�ȶǩӦۥ��ˬO�H���ʿ�Ǥ覡�@�N�uŧ�ӨӡA�]���ѽɽu�� DNA�i�l���ӽؤ��ۿ˺ح�C�Y�N�ӷ~�i�����إ��ަ^���˷��A�����Ƚt�ּۤƴX�Y�إ��ޡA�u�n���R�o�X�Y�إ��ޡA�k���X�㦳�U���S�w�H����DNA�A�N�i�Φ��ɽu��DNA�����νs�X (Barcode)�A�H�ϧO�ӷ~�i�����ަסC�ثe�x�W�ؽަ��@�Ӳɽu���]���X�A�J�ޤ]�����ު��ɽu���]���X�A�H�Τ������]�B�h����]�B���]�B�y���J�հ�]�B�ذ��ư�]�B��q��]���V�����]���X�C�ӫ~�y�q�W�A�@�Ӱӫ~�q�`�֦��ƭӽs�X�νX���A���ǽX���ȧ@���~��ި�Ϊ��y�z�ΡC�Ҧp�Ť��|����t�A�@��Ѭ���i�f�Ť��N���G�N�P�ɾ֦��n�����BNAAB�s�X�B�����X�B���W�B�ꤺ��n���������A�]����]���X���ظm�A���|�P�y���A�A���~�l�ܺz�t�Ρz�ĥΪ�EAN/UCC�X���ҽĬ�C�ӷ~�ާ@�ɡA�ѩ��]���X�s�X��ëD�C�@�ʪ����Ŧ׳J�ӫ~���ݤ����˴��A�Ӷȧ@���ò��ӫ~������~�ޡA�ǥH�O�ٮ��O�̤ΰϹj���~�C
�ʪ��ɽu���]���X�O�H���ʿ�Ǥ覡�@�N�uŧ�ӨӡA�b�x�W�ϥͪ��ʪ��|�����˪��ɽu���]���X�A�]�������w�Dzɽu��DNA�ǦC�|�o�{�a�ڮt���ΦP�M����t���C�p���@�ӡA�겣�Ŧ׳J����S�w���ɽu���]���X�C�ʪ��ɽu���]�����w�ǥ]�A12S rRNA�B16S rRNA�B22��tRNA�B13�ӳJ�ս��]��D���ϡC13�ӳJ�ս��]�]�AND1�BND2�BCOI�BCOII�BATPase8�BATPase6�BCOIII�BND3�BND4L�BND4�BND5�BND6��Cytb�C
�b�թ��C���O�դh��2003��2005�~���A��b�թ��`�ҹ}�i���ŷ紵�B���J�L�B�����J�B�L�J�L�B���α��s�����ӫ~�ؽءA�p��111�Y�ް��i��ɽu�� DNA�� D����(D-loop) 1175 bp�ǦC�����w�ǡA�o��ɽu�� DNA D-loop�� SNP�զX�����C�ءA�B�b�ǦC��m711�ᦳ5��-CGTGCGTACA 10 bp�����ƧǦC�A���ƼƬ�17-27�Ӥ����C�ڭ̥i�NSNP����������苷��A�BT�BC��G�A�P���ƧǦC�ƥؤƦ��u���P�ťաA���Ӥ@�w���s�X�W�h�զX��DNA���νs�X�C�X�i�ܨ�L�b�V�~�ؤ��ɽu�� DNA��L�ϰ�w�ǡA�p�ӭM���c���酶I ��]�]Cytochrome C Oxidase subunit 1, COI�^���A�βɽu�� DNA�����w�ǡC���ӭY�A�̳J�ս��]��ǥ�ND1�BND2�BCOI�BCOII�BATPase8�BATPase6�BCOIII�BND3�BND4L�BND4�BND5�BND6��Cytb���i��ħǦC�A�Y�i�������@�s�X��Ʈw�A�ӥѦ���Ʈw���X�ҫإߪ�DNA���νs�X�N�A�i�c���ӷ~�Ͳ����ަײ��~���f�V�l�ܨt�ΡA�H�w�����~���B��B�y�q�B�P��L�{���i��o�ͪ���_�欰�A�N���Q���U�겣�ަת��~�P�ơC
�O�ة�b�թҪὬ�����x�W�¤s�ϡA��ɽu��DNA�ǦC�g��̪߫F��ޤj�DZi�q�q�դh��2005�~�w�ǫ�A�����ǦC��16,641 bp�A�]�A12S rRNA�B16S rRNA�B22��tRNA�B13�ӳJ�ս��]��D���ϡC�x�W�¤s�ϲɽu��DNA�����ǦC�t33.53% A�B27.32% T�B13.14% G�P26.01% C�A�䤤GC��Ҭ�39.15%�C�x�W�¤s�ϸ��q�j�Q����s�ϡ]16,640 bp�^�h�@���P��A���16S rRNA (��1092��2664�P���m)�CD-���Ϥ���15811��15964��m�A�x�W�¤s�ϻP����s�϶���6�ӳ�@�P���ܲ��I(SNP)���t���C��~�زɽu��DNA��13�ӳJ�ս��]��SNP�t���g��Ķ���i��ħǦC�A�̳J�ս��]��ǥ�ND1�BND2�BCOI�BCOII�BATPase8�BATPase6�BCOIII�BND3�BND4L�BND4�BND5�BND6��Cytb���i��ħǦC���t�����O��0.9�B5.5�B0.4�B0.0�B0.0�B3.1�B0.4�B0.0�B0.0�B0.2�B5.9�B2.3�P0.2%�A�����t����1.5%�F�䤤�Ȧ�COII�BATPase8�BND3�PND4L ��4�ӳJ�ս��]�i��ħǦC�����ۦP�C�i��ħǦC�t��1.5%���s�b�A��ܨ�~�ت��h��SNP�O�\��ʰ�]�ծt���C�i�@�B�����i��ħǦC�t�����p�x�W�¤s�ϻP�q�j�Q����s�Ϥ���ǶZ����0.018089�A�q�j�Q����s�ϻP��Z�����Navajo-Churro���Ϥ���ǶZ����0.049188�A�x�W�¤s�ϻP��Z�����Navajo-Churro���Ϥ���ǶZ����0.047170�A��ܥx�W�¤s�ϰ_���ɥN����q�j�Q����s�ϡC�x�W�¤s�ϲɽu��DNA�����ǦC��SNP��T�N�i�ѥ��b�巽�l�ܤΫإߺإ��Ϫ���]���X�A�@���ئϥͲ��i������ǼаO�ΰ�]��ت��������ѡC
�ӭM�֤����V����A�]�A��V����ΩʬV����C�ʬV����q�`�Ȥ@��A���Űʪ��OXY�A�����OZW�A�W���ʧO�M�w��]�C�a��(�V����2n=38)�������ǦW�����ݹ��ƽ�(Sus scrofa domestic)�A�O�Ѽڨȳ���(�V����2n=36)�t�ƦӨӡA�k�ݩ�ެ�(Suidae)�B�ިȥ�(Suiformes)�M�����(Artiodactyla)�C�@�Ӱʪ��s�Q�����u�~�ءv���@�P�w�q���o�s�ʪ��Ӧۤ@�Ӧ@�P�����A�㦳�i�Ϥ����S�ʡC�x�W�a�Ͻب̤������魫�j�p�A�i����100���筫���j�髬�آw�ŷ紵�B���J�L�B�����J�P�L�J�L�F50���筫�����髬�آw���F�M20���筫���p�髬�آw�����C�N��38���V����W����ǰ�]�Ҧb��mø�����a�Ϣw�u�ް�]���Сv�A�A�A�ѳo�\�h��]�b�ް��ʪ��W�ҧ�t���\��A�i��d���u���]���ؽޡC�Q�ΰ�]�˴��ӿ�ܺؽޡA�ḛ̀�]���Ӱt�ءA���|�X�Ӫ��ؽN�O��]��ت��u���ޡv�C�i�@�B�w��Ť�(�V����2n=60)�B�s��(�V����2n=60)����(�V����2n=78)����ǰ�]�Ҧb��mø������]���СA�ḛ̀�]���Ӱt�ءA�i��d���u���]���دb�V�C
�|�B�دb�V�Ͳ��i���ΰ�]�n��
�b���k�ĤQ�C���u�D�����o�����ˬd������دb�V�~�̤��دb�V�B�ط��B�]�ơB��εn���Φ��������A�دb�V�~�̵L�����z�Ѥ��o�W�סB���`�Ωڵ��C�دb�V�κط��g�e���ˬd������A�o�{���k�w�ǬV�f�ο�ǩʯe�f�̡A���o���c�ޥΡC�v�C�ĤQ�C���O��]�n���B��]���X�ΰ�]��ت��k���C
|
�������k |
���q�ؼ� |
�w���įq |
|
�O�� |
�s�γ~ |
�a��� |
|
�|�� |
�s�~�ث~�t |
���~�� |
|
��]��� |
�s���� |
��ڤ� |
�x�W�دb�V�X�f�O�b����ު����P���~�A�ǥ�DNA���νs�X�BIT�N�B���ݺʴ��t�Τ�KM�t�ΡA��X���Φb�دb�V�c�ީʯ��˩w�Ϋ~����i���W�A�Ѻدb�V�M�سJ�X�f�����a���ѽX�ΡCSNP��]���X�O�HATGC���N�X�AUTR��]���X�H2��9�����ƧǦC�ơA�p���A�ǥѰӫ~��P14�X��A�i�ĥ�10��SNP�M3��UTR�ӥN���ӧO�دb�V����A�@�ӽX�ѰϤ��b�V���اO�A�զX�᪺���νs�X��536,870,912�ӡA��N���x�W���دb�V�`�ơC�x�W�إ��b�V�|���@�Ӱ�]���X����]��ǯS�ʡA��N�]�|�����˪���]���X�A�G�@�M��a�κدb�V�Ͳ��i���ΰ�]���X��X���Ψt�ΡA�i��U�x�W�دb�V�Ͳ��~�̤ΥX�f�~�̦�P�O�W�u��b�V�C�ثe�x�W�ؽޡB�ؤ��κئϦ���εn���B�ʯ��˩w�M��]�˴��A�Ӻ����κس��w���������A���n�B���Z�κبߤ]���~�t��|�A�]���x�W�دb�V�����㪺�Ͳ��i���C�b������Ұw��O�W�֤ߺظs���u�����ӫإߪ��b�VBAC��]�w��2005�~�������ޤ��ϳ������n�ZDNA�w�A�|��SNP��]���X��UTR��]���X�C��2006�~�������������Ǫ��ɽu���]���X�w�A�|����]���X�������Ѻدb�V����Τѥͼ��Ѫ��C
�دb�V���~�O�b�����~�������Y�A�b�����i��a�L�������دb�V���~�A�����دb�V���~�]�~��ϯb���~����g��C�ڰ�۰�~�ءA�O���ʯ��A��ؤΨ|�إX�s�~�ؤηs�~�t�h�~�A��~�ӧ�i�@�B���Τ��l��ǾǤu��Ӱ��|�x�W�دb�V�A�o�ذ�]��اN�]�A��]�˴��B�ʯ�����ΰt�ص������T�j���A���ԲӪ��Ͳ��i���ΰ�]�n����ơA�]���A�B�ΰ�]��اN�O�دb�V�ʯ��}���s�ͶաC�ŷ紵�B���J�L�M�����J�ة�����ؽ��P�ؽ��˩w���A���|���C�Y�ؽު���εn���ҩ��ѡC�ؽ��˩w�����˥X�⪺�C�Y�ާ��i���]�n���A�˴��ް_���`������](AA, AB, BB)�B����ƧZ�ƶq���h����](MM, MN, NN)�B�H�μv�T��٦פ����תհ�n���G�����](HH6, HL5, HL4, HL3, LL2, LL1, LL0)�A�o�T�ذ�]�˴��ODNA�ǦC�����I�P��t���C�ؽ���z�ؽޤ��ɪ��C�Y�ާ��i���]�n���C�b�թҡB�����b���|�Ϋ̬�j���ɥx�W�Ϻؽ��~��|�U�ؽ����ؽޤ��ɷ|����X��ؽޡA�i�氪���]�˴��P��]���ҡA���u��ؽު�����ǯS�ⴶ�M���Ω���A�����ަפ��~��P���[���ȡC���]�O��Ŧ�תջĵ��X�J�հ�](Heart fatty acid-binding protein, H-FABP)�A���ު��Ĥ����V����W�A���]�P�٦פ��תէt�q(�j�z�ۯ�)�����A���]�P�פ��X��סB�h�ĩʻP�}�n�����e�������C���]����Hh�BAa�BDd���T���˴����Ӽv�T��A���]���N���O��ӭ^��r���M�@�ӼƦr�F�^��r�� HH=�����]���զX�BHL=�����]���զX �B LL=�C���]���զX�F�Ʀr��0��6�A���ܲĤ����V�������V����W���X�Ӽv�T�誺���Q�����](H�Ba�Bd)�F�Ʀr6����6�Ӧ��Q�����](HHaadd)�A�G�k����HH6�F�Ʀr3��5����3�B4��5�Ӧ��Q�����]�A�G�k����HL3�BHL4�BHL5�F�Ʀr0��2����0�B1��2�Ӧ��Q�����]�A�G�k����LL0�BLL1�BLL2�C
���ӿ�Ǩ|�ؤu�@�W�A�ؽޡB�ؤ��κئϦ���εn���B�ʯ��˩w�M��]�˴��A�Ӻ����κس��w���������A���n�B���Z�κبߤ]���~�t��|�A�]���x�W�دb�V�����㪺�Ͳ��i���C�ثe�x�W�Ϻؽި�|���ؽ��������z�L��|���f���W�ؽޤ��ɷ|�C�@���Կ�ް��t�СA�ݦW��T�{��}�l�i��Ħ�ʧ@�A5~7���ֽް��Υ��˴���]�������b�ұĦ�i���]���ҡCL�PY�~�إ����O��]AA���ް��~�i�ѥ[�h����]�Φ��]���ҡAD�~�ؤ��\��]AA����AB���C��]���ҦX��ؽ�DNA�˫~�~�˴����]�C�N���]�˴����G�i���ؽި�|�ؽ��A����D��A�X�ؽްѥ[���A��Ƶn���ܥx�W�b���ح��T��(www.angrin.tlri.gov.tw )�A�ؽީ��e�A�����������]HH��HL����]���Ҭy�{�C�b�թҺ����i�[��]��حp�e��2005�~�w�����G�ؽީ�����A�����J�a�x�̰�����70�U���B�ŷ紵�a�x�̰�����52�U���B���J�L�a�x�̰�����14�U���C�~�ӡA�o�ذ�]�n���ؤ��ޤ���N����`���A�����J�~�س̰��w��466�U�������B�ŷ紵�~�س̰���192�U�������B���J�L�~�س̰���64�U�������A�����C�Y�ؤ��ަ~����30�U���C�x�W�Ϻؽ��~��|�|�����`�@�}�i700�Y�u�褽�ްѥ[�b�թҺ����i�[��]��حp�e�A���p�C�~��2�����H�W���ؽP��ȡC
�ŷ~�o�i�W�A�]���u��������|�u�@�A�ù��y�A���q�Ť����૬���ؤ����A�ǥH���Ѱꤺ�u�誺���Ťκؤ��A��N�����|�X����Ǥ��ܪF�n�Ȱ�a�C��F�|�A�~�e���|�b������ҩ�1996�~�}�l�ʱ���~�i�f���Ť��N���G��ǰ�]�I���A�ì�o�����c�ް�]���ֳt�˴���k�A�P�B�˴����Яg�]DUMPS�^�B�O�ڲy�H�O�ʥ��g�]BLAD�^�Υ��i�įg�]CITR�^�� 3 ���c�ް�]�C���Яg��]�|�ϥ���������Z�����ۧɡB�t�ب����v�U�� 30�H ���h�� 40��60 �ѭL�ভ�����`�F���i�įg��]�|�ϥX�ͤ@�g���J���B�藍í�B�����B���z���`�F�O�ڲy�H�O�ʥ��g��]�|�� 2 �g�� 8 ���֥J���զ�y�L�h�B�ͪ��w�C�B�Y���U�g���Ӧ��A�è��h��������L�u�y���v�W�[�C��X�H�W�ҥܪ����а�]�B�O�ڲy�H�O�ʥ���]�B���i�İ�]�B�۩ʰ�]�ζ��ʰ�]���o�����c�ް�]�˴��N�A�i���Ħa�����U�ؤ����i��k���c�ް�]�E�_�A�إ��u�}�����s�C
���B��]��ؤα��X�ƪ�����
�]���g�T���y�ơB�ۥѤơA�x�W�A�~�����q�u�~�ا�}�v�B�u�~��O�ҡv�Ρu�~�P��P�v�}���v���Q��A�[�j�зs��o�B��ڦ�P�Υ��~�Ƹg��A�غc�A�~�[����t�A�ɯŬ���o�ƪ��@�Ų��~�B�����[���Ȫ����~�[�u�~�A�Ψ㦳�S�⪺�A�~�A�ëإ߹A���~�åͦw���{�Ҩ�סA�����O�̯���w�ߨɥΦw�����A���~�A��y�x�W�A�~���s�槽�A���T�x�W�A���~�~�P�A���x�W�A�~�b�s�@�����T�O�v���u�աA��������o�i�����P���~�C�H�W�o�q�ܬO����2004�~2��25����ɹA�e�|���D�e���s�b�ߪk�|�Ĥ����Ĥ��|���g�٤ί�e���|���X�~�ȳ��i�����\�C

�x�W�ѵM�귽�����A�H�۸g�T�ۥѤƻP��ڤƵ{�ת������A�x�W�b���~�w���{���ȡB�g��IJv�B�����v�������ҫO�@�����D�C�ަײ��Ȧb2002�~��492.3�����A�C�H�C�~��49���窺�ަץi���O�C�ަײ��ȬO�b���~���Ȫ�46.8%(492.3����/1051.9����)�A�Φa��3,734�����A�C�������~���Ȧ�1,318�U���F����b���~�ӬݡA2002�~�C�������~���Ȧ�925�U���C����}�~�V�A�����q�u�~�ا�}�v�B�u�~��O�ҡv�B�u�~�P�еo�v�B�u�~����ơv�Ρu�~�w�z�v�}���v���Q��C���ɶDzίb���~�N����O�A�ͪ��N�Y��������N�C�]���A�b���~��]��اN���o�i�A�O�x�W���ӯb���~����o�i�����ǡF���P�ɡA�����ͪ��N����s�зs�o�O���{�@�ɩʪ��E�P�v���C�ҥH�A��]��اN��o�P���ήIJv������A�O�T�O�b���~��ŦX��a�ݨD���ߤ@�~�|�C
�x�W�b2003�~�O���y��21�W���i�ް�a�A�Ӭ���ƦW��2�W�B�[���j�ƦW��12�W�B�饻�ƦW��16�W�C�x�W�ؽޤ��}�ƮIJv(�}��/�W��)��2.19�A�Ӭ��[�ؽު��}�ƮIJv��2.67�A��ܰ�]��ث�|�����u�����~�إX�{�C�]���A���x�W�u��ޫ~�إX�f�A�óz�L��ڦ�P�X�@�覡�A�i�o�i�X�A�X���w�ڦ�(��3�W�i�ް�)�B�����(��9�W�i�ް�)�B�ڬw��Z��(��5�W�i�ް�)�ΨȬw���a�a�Ϫ��V�n(��6�W�i�ް�)�B�L��(��8�W�i�ް�)�B���(��14�W�i�ް�)���ءC
|
���� �x�W�b1961�B1991�M2001�~���a�b�V�~���b�i�Y�ƻP�~�`���� |
||||||
|
1961�~ |
1991�~ |
2001�~ |
||||
|
�اO |
�Y�� |
�اO |
�Y�� |
�اO |
�Y�� |
�Φa���� |
|
���a�� |
178,394 |
�� |
10,089,137 |
�� |
7,164,605 |
3,734 |
|
��v�� |
13,631 |
�@ |
�@ |
�@ |
�@ |
�@ |
|
���ؽ� |
2,913,451 |
�@ |
�@ |
�@ |
�@ |
�@ |
|
���� |
318,162 |
���� |
18,618 |
���� |
6,531 |
�@ |
|
���������ؤ� |
96,679 |
���������ؤ� |
33,699 |
���������ؤ� |
12,258 |
�@ |
|
�Ť� |
3,301 |
�Ť� |
100,539 |
�Ť� |
133,718 |
�@ |
|
�@ |
�@ |
�@ |
�@ |
�� |
�@ |
1,504 |
|
�s�� |
136,297 |
�s�� |
175,346 |
�s�� |
184,717 |
�@ |
|
���� |
703 |
���� |
419 |
�@ |
�@ |
�@ |
|
�@ |
�@ |
�Ŧ� |
39,600 |
�Ŧ� |
99,388 |
�@ |
|
�@ |
�@ |
�@ |
�@ |
�� |
�@ |
531 |
|
�� |
222 |
�� |
750 |
�� |
805 |
�@ |
|
�� |
7,914,941 |
�� |
79,120,000 |
�� |
117,310,000 |
3,671 |
|
�n |
3,910,794 |
�n |
10,661,000 |
�n |
10,104,000 |
1,202 |
|
�Z |
1,474,588 |
�Z |
1,931,000 |
�Z |
2,613,000 |
408 |
|
���� |
328,677 |
���� |
284,000 |
���� |
235,000 |
38 |
|
�� |
649 |
�� |
32,842 |
�� |
21,189 |
236 |
|
�@ |
�@ |
�� |
139,050 |
�� |
41,523 |
�@ |
|
�~�`���� |
4.9���� |
�@ |
895.6���� |
�@ |
1,051.9���� |
�@ |
|
���A�~�`���� |
20.93% |
�@ |
37.55% |
�@ |
30.02% |
�@ |
|
�@ |
�@ |
�@ |
�@ |
�@ |
�b���Φa���� |
11,370 |
�����ӬݡA�x�W���b(�ޤ���)�V(���n�Z)�|����t�|�i�B�����P��o�ץ��s�������ͤθ�T�N�A���U�دb�V�ʯ��i�P���g�a�b�V���Ų�w�A���ѧ@������|���̾ڻP�}�o�㥻�g�ƯS�ʷs�~�t�������C�s�~�ث~�t�b�V�ʯ��|�N����o�P���s�A���i�氪�Ͳ��IJv�|�ؤ�k��s�A�ñ��s�������دb�V�~�̥H���ɨ䪪���g��IJv�F����|�ʪ��N�H�����L�k�i��̬��n�C�ô¦V�\��ʰ�]���s�A�إ�EST(��]���{�ǦC)��Ʈw�A���X�ʪ���ƻP�{����]��Ʈw�A����쥻�g�b���ح�S�����g�٩ʰ�]�A�i�@�B���Q��\��P�ձ�����A�äޤJ�|�رڸs�H�|�����v���O���~�t�C
�g�٨|�ؤW�A�̲��~�O��X�㭫�n�g�ٻ��Ȥ�����دb�V��Ǹ귽�A�����j����a�Ů֤ߺدb�V�ڸs�A�[�t����دb�V�P�B��}�}�B�P�������~�v���O�C�[�j����دb�V�|�ظ�Ʈw�P��T��������K�ʡA�F���u�}�دb�V���n�g�٩ʪ���ڵ���P�ꤺ�ƦW����ΥؼСC���X���ͧN�P�ƶq��Ǿǭ�z���اN�����ΡA�إ߿�ǼаO���U��ިt�ΡA�B�v�B�N���n��ǼаO�ǤJ�دb�V�n���t�ΡA�غc���㤧�دb�V��Ǩ|�ظ�T�C
��]���X�b��a�κدb�V�Ͳ��i�����g�ٮįq�ΰ�]���X��X���Ψt�Τ䴩�X�f���x�W�دb�V�Ψ��Dz��~�Ͳ��i����t�A�i�����Ͳ��i���ΰ�]���X�]�֡A�T�O�x�W�دb�V�~�ؤΫ~�誺���a���Ѫ��[���ȡC��]���X�̲ץi�PRFID�����s�X�N���X�A�ר�O�㦳�ѧO�\����s�X�C���ΰʪ��}�i���H�Ͳ��i���ΰ�]���X��X���Ψt�κz�A���~��P�A���XRFID�A�i�קKRFID�Q�ʪ��r�a��ή��O�̦Y�쪺���I�F�P�ɦ���]���X�����b�S�ʡA�b���~�L�k�Q��_�C
�ѦҤ��m
Edwards SV and AC Wilson. 1990. Phylogenetically informative length polymorphism and sequence variability in mitochondrial DNA of Australian songbirds (Pomatostomus) Genetics 1990. 126: 695-711.
Hebert PD, S Ratnasingham, and JR deWaard. 2003. Barcoding animal life: cytochrome c oxidase subunit 1 divergences among closely related species Proc Biol Sci 2003. 270: S96-S99.
Hebert PD. 2003. Biological identifications through DNA barcodes Proc Biol Sci. 2003. 270: 313-321.
Hebert PDN. 2004. Identification of Birds through DNA Barcodes PLoS Biol 2004. 2: e312.
Mayr E. 1942. Systematics and the origin of species from the viewpoint of a zoologist New York: Columbia University Press 1942.
��F�|�A�~�e���|�b������ҤE�Q���~�Ĥ@�u�dzN��Q�|(2006/03/14)